

Adoro guardare al passato e rendermi conto di quanto tutto quello che utilizzo oggi, almeno la. Aggiornata delle cose che utilizzo quotidianamente in casa mia, viene da invenzioni molto antiche. Cominciamo con gli antichi romani, i romani ci hanno aiutato veramente tanto dal punto di vista di costruzioni prima di tutto. Come vediamo in Italia è pieno di costruzioni romaniche che ancora oggi si sorreggono perfettamente.
Le costruzioni romaniche, sono in tutta europa e siamo riusciti a portare anche in Inghilterra, cioè in tutti i posti che sono stati conquistati dai romani, e le fogne sono state un’ottima invenzione soprattutto dal punto di vista igienico, che hanno permesso anche lo smantellamento dei rifiuti. La maggioranza dei sistemi di fognatura in tutta Europa deriva ancora dalla creazione e dall’invenzione di quella che è stata quella dei romani.

Chiudi gli occhi e immaginati un mondo con macchine che vanno senza petrolio, macchine elettriche o a idrogeno, perché in fondo non c’è bisogno di andare veloce, mai, non ha senso, e poi il vecchio detto del chi va piano, va lontano funziona sempre. Per prima cosa, se le macchine avessero un limite di velocità, che io metterei a 90 nelle autostrade e a 40 max nei paesini, sicuramente dimezzeremmo gli incidenti e tutto quello che ne consegue.
Inoltre le macchine vanno così veloce perché utilizziamo il petrolio per farle funzionare, tutt’altro che necessario. Come vediamo,esistono macchine elettriche, a idrogeno e metano, per cui non c’è bisogno del petrolio,lo fanno diventare necessario ma non lo è! Odio uscire di casa e fare fatica a respirare per lo smog, odio stare un coda sulle autostrade e respirare veleno,m perché non riesco a vederne la necessità.

Sono sempre stata convinta che la Scienza può aiutarci in molti modi diversi, perché il suo scopo sarebbe quello di capire come funziona la natura e riuscire a trarne il massimo beneficio. Nello stesso tempo è molto importante capire come lavorare con la natura senza distruggerla, come stiamo facendo. Se ci pensiamo bene, la natura rappresenta la forza motrice della terra, noi siamo solo ospiti, ancor più degli animali, che contribuiscono alla natura in maniera del tutto naturale.
Noi, esseri umani no, siamo gli unici che producono inquinamento che la natura non riesce a processare, e siamo gli unici con libero arbitrio e ragione, almeno così pensiamo. In realtà, anche la piante hanno una loro energia e hanno così tante proprietà che i medicinali delle farmacie ti faranno dubitare di alcuni passi falsi della scienza, che diventa capitalismo, come tutto il resto. Beh, per prima cosa, lo sapevi che la maggioranza dei medicinali vengono dalla sintetizzazione di sostanze naturali?
Che bisogno c’è di creare qualcosa chimicamente che la natura può darti naturalmente? La risposta non è difficile, basta pensare a quante ingiustizie ci sono nel mondo, per questa ragione, non mi sorprendo quando le persone non sanno che possono addormentarsi solo con la camomilla e con la valeriana e che non hanno bisogno di sonniferi. Il discorso delle piante diventa più complicato solo perché le piante hanno moltissime proprietà, ogni singola pianta o arbusto ha più di una proprietà che può essere relativa ai benefici che può apportare al nostro fisico.
Le piante hanno arbusto, foglie e radici, per non dimenticare la fioritura, tutti questi aspetti e passaggi sono di fondamentale importanza e permettono di poter davvero curare anche i piccoli disturbi, se le conosciamo bene. Conosco due farmaciste che sono andate in Umbria e hanno comprato un appezzamento di terreno nel quale poter piantare le piante e studiare le loro proprietà, in maniera da assorbire le loro qualità, e trasformarle in modo da farle risultare benefiche per noi!
Le piante sono una risorsa incredibile, anche quando vai a passeggiare nei parti, ti sorprenderesti nel vedere quante piante puoi utilizzare, per non parlare degli oli essenziali e dei fiori di Bach, nei quali parlerò in un altro post. La natura e la nostra chiave per capire meglio la terra, e chissà perché l’uomo e la sua ragione sforzano sempre troppo la serratura!

Sono andata a cena da amici l’altro ieri, e ho notato che nella loro credenze c’erano una miriade di padelle, mi sono chiesta anche cosa servissero tutte quelle padelle così diverse, anche perché io nella mia cucina ne ho tre tipi classici, la casseruola media, una piccola per il latte e una pentola di alluminio antiaderente, ho notato che il mio amico stava cucinando nel coccio, stava preparando la zuppa di cipolle. Parlando con lui, ho saputo che il coccio è un tipo di ceramica che si resta molto bene per la cottura di zuppe o minestre proprio per la capacità di questo materiale di condurre il calore.
Si tratta davvero di uno dei migliori materiali per questo tipo dai pietanze, mi hanno raccontato, e io mi sono ricordata dei minestroni di casa mia, e la pentola di coccio di mia madre. In realtà, anche la porcellana si presta molto bene alla cottura di verdure, ma sembra molto più delicata della terracotta, il coccio. E poi in quella credenza,ma casa dei miei amici, noto una padella in pietra, molto bella, e scopro che si tratta di i una pietra metamorfica che si utilizza per cucinare carne e pesce, e il rupi saltato sembra migliore di quando si utilizzano le padelle classiche in alluminio.
Intendi dire, che i miei amici hanno cucinato in quella padella e che mi è sembrata la verne più buona che io abbia mai provato. Il giorno dopo sono andata dal negozio per casalinghi vicino casa e ho acquistato la mia padella in pietra ollare per la mia carne e pesce, per poi scoprire che per pulirla non avevo neanche bisogno del detersivo e ti permette di cucinare senza grassi! Ovviamente coccio, porcellana e pietra non sono così abbordabili, come prezzo, insomma non era davvero economica, quindi puoi cucinare anche con le altre padelle antiaderenti, basta solo sapersi prendere ben cura di loro!
Le antiaderenti e come pulirle!
Le padelle in alluminio sono tra le più economiche sul mercato, ma non altrettanto qualitative, il problema e che si rovinano molto più facilmente rispetto a quelle con materiali naturali,per cui potrebbe capitare di graffiarle, se succede non utilizzarla più, anche perché la padella diventa cancerogena, fa male alla salute,perché la polverina antiaderente che graffi è velenosa.
Quindi quando utilizzi la tua padella in alluminò antiaderente, assicurati di non utilizzare posate di metallo o raschietti per pulirla, al contrario, se di essere essere particolarmente incrostata di grasso, lasciala in acqua bollente e detersivo, o aceto e sale per tutta la notte, il mattino riuscirai a pulirla che è una meraviglia e punì questo modo preservi anche il suo stato!

Quando la mia migliore amica mi ha confidato che stava diventando mamma, ho subito pensato a quanto passa veloce il tempo. Una delle cose che mi ha sempre fatto paura, la velocità del tempo. Il tempo ancora e ancora.Torna come unico protagonista nella nostra vita, il tempo e’ vita e la vita e’ nel tempo. Gli uomini fanno riferimento al tempo in continuazione.Ogni azione dell’uomo e’ soggetta al tempo e molti tra i discorsi umani si riferiscono allo stesso. Ci sono delle volte, durante le quali desideriamo che si fermi, altre nelle quali desideriamo che passi il più’ velocemente possibile. Il mio rapporto con il tempo e’ cominciato da poco.
Il problema sta nel riconoscere e comprendere la sua silenziosa esistenza. Si e’ sempre troppo impegnati a crescere per pensare a lui, ma poi ti svegli un giorno con la consapevolezza di aver trascorso un quarto di secolo su questa terra e non te ne sei nemmeno accorta. Non ti sei resa conto che sono passate decine di anni, migliaia di giorni, milioni di ore e miliardi di secondi, dalla tua nascita. Mi spiego meglio. Il tempo di per se’ rappresenta l’invisibile condizione della vita.Condizione necessaria e sufficiente della vita in quanto ogni forma di vita e’ soggetta al tempo.
Cosa c’entra con la mia amica? Che siamo diventate grandi e lei mi ha chiesto di accompagnarla a comprare un passeggino…ma quale? Io ero abituata al passeggino classico a quattro ruote, tipo quello della chicco,una seduta molto semplice e con al massimo un ripiano basso per appoggiare quello che si vuole, e invece oggi scopro che la scelta del passeggino è tra le più complicate per i genitori, perché si tratta di un universo di scelte, e nella maggioranza dei casi l’investimento supera le aspettative, e ci credo, con quello che costano!
La tecnologia della leggerezza!
Ovviamente la Scienza non poteva che andare incontro alle necessità delle mamme e così hanno creato dei passeggini super leggeri e super pratici. Siamo andate in un mega rivenditore e abbiamo scoperto dei modelli che si aprono e chiudono con una sola mano, che sono in grado di andare su asfalto e prato,con delle ruote davvero giganti.
Ma non è tutto, se pensate che le ruote siano sempre 4 vi sbagliate, oggi potete trovare anche versioni a tre ruote, e modelli multifunzione,in grado di trasformarsi in modelli diversi! Ovviamente i prezzi qui, cambiano notevolmente,ma si sa stai investendo per il tuo piccolo e ne varrà sempre la pena

Sono diventata una vera salutista e presto sempre molta attenzione a quello che mangio, quindi ho pensato di cominciare a cucinare con l’uso del vapore, anche se qualcuno mi ha avvertito che molte volte il vapore toglie il sapore al cibo. Non sono d’accordo, del tutto. Ovviamente prima di comprare una vaporiera pensa bene ai modelli disponibili in commercio, così saprai di quale hai davvero bisogno. Ci sono un sacco di modelli a basso costo in giro, e le vaporiere hanno davvero un sacco di svantaggi; per prima cosa, si di uno di quegli oggetti da cucina, che spesso finiscono relegati e messi nella parte posteriore della credenza della cucina e dimenticati. Sbagliato.
Devi sapere che la cottura a vapore mantiene il numero maggiore delle vitamine e nutrienti negli alimenti rispetto ad altri metodi di cottura come la bollitura, dove i nutrienti vengono portati via dall’acqua. Tuttavia, mentre guardando attraverso Il contenitore trasparente le pile di verdure nutrienti, può farti sentire bene, devi anche sapere che un vapore elettrico non è in realtà il modo più sano di cucinare. Il meglio del meglio è il forno a vapore, perché mantiene più sostanze nutritive del cibo; inoltre è anche più veloce e utilizza temperature più basse e meno acqua.
Un cuoco organizzato può preparare tutto il cibo per la cottura a vapore in anticipo, e basta aggiungere i cestelli per la cottura a vapore quando è il momento di iniziare la cottura. Questo potrebbe essere un grande vantaggio in occasioni come il giorno di Natale, in cui un piano cottura completo di pentole di germogli, carote e sugo può causare un sacco di confusione.
La maggior parte dei libretti di istruzioni consigliano di non utilizzare condimento o qualsiasi altro liquido diverso dall’acqua nel processo di cottura, anche se questo non dovrebbe impedirvi di vivacizzare le cose, aggiungendo erbe fresche sopra il vostro cibo fumante. Se ti organizzi nel modo giusto, potrai portare un intero pasto insieme e lasciare il piano di cottura libero per la padella del sugo, una buona vaporiera ti aiuterà se tu rispetterai i tempi di cottura. Con la pratica è possibile tirare fuori interi pasti da una sola vaporiera, ma come qualsiasi altro elettrodomestico da cucina, l’abilità del cuoco è in gran parte quello che porta al successo o al fallimento.
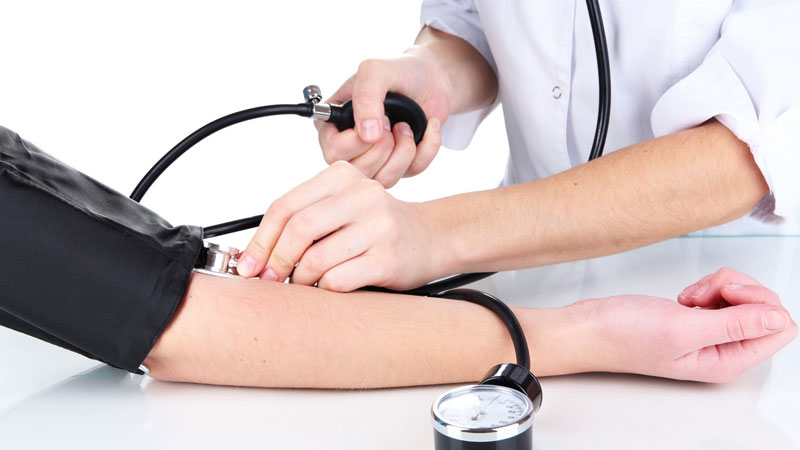
Purtroppo a casa ho qualcuno che soffre di pressione alta, e in questi casi andrebbe monitorata molto spesso, per cui invece di andare dal medico tutte le volte puoi optare per una soluzione casalinga, che ti consente di monitorare la tua pressione, tutte le volte che vuoi, e ti consiglio di segnare sempre i valori che escono, soprattutto se hai problemi di pressione, sia alta che bassa.
Se si decide di misurare la pressione sanguigna a casa, hai sicuramente bisogno di ottenere un monitor della pressione arteriosa, e devi sapere che esiste una vasta gamma di misuratori di pressione a disposizione, ma è importante essere sicuri che il monitor della pressione arteriosa che si sceglie è accurata e quello giusto per voi.
Come ho appena scritto, in commercio c’è ne sono davvero moltissimi, e sicuramente tra quelli che ho trovo, quello più semplice da utilizzare e quello digitale. Cerca di sceglierne uno che misura la pressione sanguigna dal braccio, piuttosto che dal polso o dalle dita, perché il misuratore pressione a braccio di solito danno i risultati più accurati e coerenti. Ciò significa che il monitor digitale è passato attraverso una serie di test per assicurarsi che dà risultati che voi e il vostro medico potete considerare accurati, o i più accurati possibili.
Le caratteristiche da conoscere!
Un monitor della pressione arteriosa superiore del braccio sarà disponibile con un bracciale che è necessario avvolgere attorno al braccio. Se si utilizza un bracciale che è la dimensione sbagliata per te, la vostra lettura della pressione sanguigna non sarà corretta. Misura intorno al vostro braccio a metà tra la spalla e il gomito, e scegli la dimensione del bracciale, la maggior parte dei monitor della pressione del sangue a casa sarà disponibile con un bracciale di medie dimensioni,ma se Ne avete bisogno potete tranquillamente ordinare un bracciale di dimensioni diverse separatamente.mobbiamnete il prezzo di questi accessori varia notevolmente.
E questo di solito dipende dal numero di funzioni extra che il monitor digitale ha, come una memoria incorporata, per esempio. Tutto ciò che serve per misurare correttamente la pressione del sangue è un monitor clinicamente validato, e carta e penna per registrare le vostre letture. Le caratteristiche supplementari possono essere utili, ma non sono necessari. Scegli un monitor della pressione arteriosa a casa che ti puoi permettere e che ti assicura risultati coerenti e validi.

Una macchina automatica per il pane significa che fare il pane fatto in casa è un gioco da ragazzi, perché con la maggior parte delle macchine, è solo una questione di misurare gli ingredienti nel contenitore e impostare il programma giusto. La tua prima considerazione dovrebbe essere la quantità di pane che vuoi esser in grado di poter fare a casa, perché in commercio, troverete macchine che creano pagnotte che variano da 1 a 3 chili, e molte offrono due o anche diverse opzioni di formato pagnotta nella stessa macchina.
Tenete a mente che il pane fatto da voi, con la macchina non rimane fresco più a lungo di quello comprato al supermercato; ti consigliamo di mangiare la maggior parte entro un giorno o due. Quindi, se avete una grande famiglia o mangi un sacco di pane, probabilmente una macchina che fa una pagnotta di 2 kg sarebbe sufficiente per una famiglia Medio piccola. Le macchine che offrono due o tre opzioni di formato per pagnotta sono utilissime, perché funzionano con una vasta gamma di ricette e vi daranno la possibilità di fare pagnotte dalle dimensioni diverse per le vostre esigenze o occasioni particolari.
La tecnologia dietro alle macchine per il pane!
Oggi, la maggioranza di macchine del pane fare una pagnotta orizzontale che è più simile a quello che si potrebbe comprare in un panificio. Praticamente ogni macchina per fare il pane consente un certo grado di programmabilità. I controlli più comuni consentono di scegliere la crosta chiara o scura, un impasto senza cottura, o cuocere senza impastare. Alla più complessa e costosa fascia di macchine per pane, ne troverete alcune che hanno programmi specifici per i diversi tipi di pane, che tengono conto del tipo di farina e altri ingredienti per regolarne la cottura e tempi di impasto.
Questi potrebbero includere programmi di grano integrale, senza glutine, pane francese, pane artigianale e pane dolce. E ancora altre macchine avranno programmi che consentono di utilizzare la macchina per fare più di solo pane: marmellate, torte, pasta pasta e pasta della pizza sono alcune delle cose che alcune macchine sono in grado di fare. Naturalmente, tutte queste caratteristiche in genere si traducono in un prezzo più elevati, così se stai valutando quale macchina per l’acquisto, tieni conto di quali caratteristiche pensi davvero che verranno utilizzato spesso da te, in questo modo compri quello che ti serve!

Il ricetrasmettitore fa una grande differenza quando si viaggia, app anache quando si lavora o semplicemente per piacere, questo accessori noti anche con il nome di Walkie Talkie, che in inglese significa ‘parlo e cammino’, hanno proprio questa capacità e permettono di comunicare secondo certe distanze. Quindi per esempio, per gente che lavora in cantieri o in magazzini diventano accessori indispensabili per poter comunicare, senza dover utilizzare il telefono, perché la comunicazione avviene completamente gratis.
La stessa cosa se viaggi. Lo puoi utilizzare per mantenerti in contatto se partite con tanta gente e poi di si divide o in circostanze simili a questa, quindi anche un campeggio potrebbe rappresentare un buon esempio. I bambini amano gli walkie talkie, e per questo motivo, molte aziende fanno walkie talkie specifici per loro, pulsanti facili da usare, spesso con i loro personaggi preferiti disegnati sopra.
Questi walkie-talkie diventano un modo divertente per i bambini di comunicare con la famiglia o gli amici se stanno giocando al chiuso o all’aperto, inoltre sono accessori che vanno da modelli piuttosto semplici a alcuni davvero sofisticati ed elaborati, quindi è meglio per il consumatore sapere che tipo di walkie talkie sta cercando, in modo da poterlo utilizzare anche in distanze diverse, potresti davvero sorprendenti delle nuove tecnologie.
Se vuoi comprarne uno anche tu continua a leggere!
Per tutti quelli che stanno pensano all’acquisto di un walkie talkie, ci sono alcuni fattori da considerare! Credo io abbastanza importanti per prevenire eventuali brutte sorprese. Non si tratta di un ritelefonò, quindi, rispetto anche alla zona nella quale ti trovi potresti perdere campo e comunicazione, molto più facilmente che con il telefono. Un altro fattore determinante per l’acquisto è il Budget a disposizione; quanto vorresti spendere per un walkie-Talkie e se sei, alla ricerca alla ricerca di un walkie-talkie usato o un nuovo modello di marca?
Alcuni utenti potrebbero voler funzioni extra sui loro walkie talkie, come un GPS, mentre altri utenti se la cavano meglio con il modello base walkie talkie. Ci sono walkie talkie che sono più in stile tradizionale e possono andare bene in un aggancio alla cintura, o essere trasportati, mentre alcuni sono fatti per il polso o su un auricolare per le mani libere, mentre la comunicazione gratuita che può essere molto conveniente in movimento, soprattutto se si è impegnato con attività all’aperto!

Amo dormire, amo dormire bene, e in realtà dopo il mio ultimo acquisto mi sono resa conto di quanto non mai davvero prestato attenzione al mio materasso. Beh, sono stata spesso in affitto e la casa era sempre già arredata per cui non sono mai andata ad acquistare il materasso, mentre ora nella mia nuova casetta, ho pensato anche a quello. Ovviamente non avevo idea delle tipologie e della quantità di materiali disponibili.
Ho scoperto che ci sono materassi che aiutano anche per chi ha problemi di schiena, che rispettano la forma del corpo e si modellano a misura del tuo corpo. Ho scelto un materasso in memory che mi possa aiutare a dormire nella maniera più semplice possibile, certo stipi tratta di una tipologia tra le più costose e nel caso sia troppo costosa ti consiglio il waterlily, simile al memory ma meno costoso, oppure puoi anche optare per il latex.
Dopo essere venuta a conoscenza di così tante tipologie diverse, mi sono decisa a lasciare andare una volta per tutte la mia idea di materasso con le molle. Ricordo che quando eravamo piccoli, andavamo spesso a dormire dalla nonna, avevamo la nostra cameretta, con i letti più rumorosi che io abbia mai sentito. Mi ricordo le molle di quel materasso, cigolavano appena accennavi a sederti beh, oggigiorno siamo davvero lontani da quelle tipologie, anche se credo che nella vecchia casa di mia nonna quei materassi ci siano ancora.
E dopo tutto quando penso a quel periodo, non mi viene neanche in mente di quanto dormivamo scomodi, non volevamo mai dormire in realtà. Quando ho cominciato a guardare online, ho capito che avrei trovato la soluzione giusta per me, e dopotutto sapevo anche cosa non volevo: le molle. Qualsiasi altro materiale sarebbe andato bene, ma non è proprio così! Le ultime tecnologie hanno permesso lo sviluppo di un materiale davvero perfetto per il corpo, il memory, che è davvero consigliato anche dagli ortopedici!
E se lo dovessi cambiare?
Credo di non aver mai dormito meglio, il memory aiuta e sorregge davvero tutto il mio corpo in maniera davvero ergonomica, inoltre si tratta di un investimento che arriva con garanzia, anche se non ho dubbi sulla qualità del prodotto. Se dovessi cambiare casa lo porto con me, e se dovesse invecchiare forse proverei il wateterlily, che mi hanno consigliato in tanti. Nel frattempo mi godo delle dormite davvero speciali!

Non molto tempo fa ho assistito ad una scena davvero orribile, un uomo per la strada è svenuto di fronte a me, vicino all’uomo per fortuna c’era una donna che ha tirato fuori dalla tasca dell’uomo una strana penna, che poi ho scoperto essere insulina. In effetti il diabete è un tipo di malessere sempre più frequente. Nella nostra società e questo dipende da molti fattori, stress incluso.
I cibi che mangiamo oggi sono sempre processati per cui mangiamo davvero male e molte,troppe volte assumiamo un numero di grassi e zuccheri sbagliato, perché la maggioranza di queste sostanze sono già contenute nel nostro corpo e nel cibo non processato,quindi noi sovraccarichiamo il nostro corpo di grassi e zuccheri, la maggiorana delle volte senza saperlo! Diciamo che tutto questo dipende dal sistema endocrino, che produce degli ormoni molto importanti, e il mal funzionamento di questo sistema, causa la mancanza o il sovraccarico di questi ormoni.
Le conseguenze in gergale sono molte, tra le quali il diabete, che esiste di due tipi, tipo 1 e tipo 2. In realtà i sintomi sono molto simili, e purtroppo le conseguenze non sempre piacevoli. I diabetici devono prestare attenzione a quello che mangiano perché il loro corpo non riesce a processare gli zuccheri che si accumulano nel sangue causando gravi problemi. Per questo motivo esiste l’insulina che aiuta a regolare i livelli di zucchero nel sangue dei diabetici, e l’uomo che vidi, non aveva preso l’insulina in tempo.
Perché lo zucchero nel sangue?
Succede che i valori del sangue si sballano così tanto che si perdono i sensi molto facilmente. E ti riprendi anche molto facilmente quando assumi l’insulina. Prima di quel giorno non avevo mai visto un diabetico stare male, e poi ho pensato a quando sono stat in India, e non so se lo sai ma fino a qualche anno fa, non era disponibile negli ospedali e quindi immaginati le conseguenze di questo atroce delitto.
Se vuoi saperne di più e in maniera dettagliata, sul sito degli ospedali dovresti per trovare tutte le informazioni che ti servono, compresi cosa puoi fare per prevenirlo, soprattutto se hai una storia di diabetici in famiglia. Si tratta di una malattia dell’epoca moderna, che risulta anche dallo stress continuo al quale siamo sottoposti, per questo motivo trova sempre modo e tempo per ritagliare un pezzo di giornata per te e il tuo sistema endocrini ti ringrazierà!

Sto finendo di preparare la valigia, domani parto, vado in vacanza. Come sempre riesco farci stare davvero di tutto, anche perché questo trolley mi segue davvero ovunque! Viaggiando molto spesso avevo bisogno di qualcosa di comodo ed ecco che cerco online e trovo una miriade di scelta, considerando che il mio budget è abbastanza limitato, restringo il campo della mia ricerca, e trovo proprio il modello che cerco , quello che mi segue in tutti i miei viaggi da ormai più di tre anni.
In realtà, ho sempre viaggiato con i borsoni, li ho sempre preferiti al resto e non mi sono mai fidata di quelli che mi dicevano del trolley, per me era solo troppo ingombrante e pesante, sono anche piccola, non sono una vichinga, quindi non mi sono mai immaginata con un bagaglio trolley a mano vero e proprio. Il trolley in realtà è una buona via di mezzo, che ti permette di massimizzare la capienza in poco spazio. Io ho un metodo tutto mio per mettere i vestiti in valigia, utilizzo il metodo dell’arresto lamento che ho trovato online, una vera svolta!
Quindi, mi sono guardata intorno e non sapevo se scegliere un trolley di marca in sconto, o di uno sotto marca ma di buona fabbricazione. Dopo aver letto un po’ di recensioni ho acquistato un bagaglio trolley colorato, che adoro e come ho scritto all’inizio mi segue davvero ovunque, ma ero molto indecisa sulla struttura, ho pensato scelgo un modello rigido o uno morbido?
Perché ho scelto quello rigido..
Considerando che viaggio molto spesso in Italia e che trasporto formaggi e olio ho optato per un comodissimo trolley rigido, e le mie fantastiche rotelle mi aiutano a trasportarlo dove voglio. Adesso posso viaggiare anche con la valigia stra piena, perché con le rotelle sono in grado di trasportare il mio trolley ovunque e ovviamente mi sono ricreduta sulla sua capacità di aiuto. E poi guardando indietro, ho pensato a tutte le volte che trascinano il mio borsone come una matta da aeroporto in aeroporto, quindi sono davvero soddisfatta della mia scelta.
Quanto amo quelle rotelle adesso, non saprei davvero cosa fare se si rompessero o se non le avessi! Inoltre, grazie alla sua struttura rigida posso metterci cose molto delicate, stando davvero tranquilla che tutto il contenuto rimane perfettamente intatto! E se anche tu come me hai avuto gli stessi dubbi, spero di averti schiarito le idee!

Non sono mai stato particolarmente bravo a gestire il tempo, penso che trascorre troppo in fretta e non mi sono mai preoccupato di considerare la sua presenza silenziosa, che sensazione di impotenza tutte le volte che penso al tempo. Per fortuna ci sono gli orologi, che lo segnano, che ci rendono partecipi del suo passare, e devo dire che sono una persona che controlla spesso che ora è. Non utilizzo orologi da polso, al contrario guardo sul cellulare, mi piacciono gli orologi da polso, ma non essendo mai stata abituata ad indossarla mi dimentico di metterlo.
Al contrario in casa mia ho due orologi da parete, che per qualche tempo segnavano ore diverse e nessuno dei due era quello giusto. Per un po’ di tempo in e piaciuto vivere senza sapere che ora era, almeno sapevo di non poter far riferimento agli orologi, anche se il loro design mi è sempre piaciuto. Anche quelli con pendolo o cucù, entrambi i quali si trovano a casa dei miei genitori, sono tra le tipologie che preferisco in assoluto! E ora ho sistemato uno dei due orologi di casa mia, così posso sapere che ore sono anche perché non tengo il cellulare attaccato a me. Forse il mio rapporto col tempo e cominciato da poco, ecco perché ho deciso di sistemare il mio orologio.
E con il tempo…
Ciò’ che mi spaventa in realtà’ e’ il sapere che il “ciò’ che fu” e’ immodificabile. Nella consapevolezza della libertà’ di scelta del presente e del futuro, rimane invalicabile la presa di coscienza con il tempo passato, come una saetta silenziosa. Mi spaventa la certezza di non poter modificare il passato e neanche l’ultimo centesimo di secondo che mi porta a scrivere su questo foglio. E ancora non posso cambiare il tempo immobile della sofferenza e non posso bloccare il tempo rapido della felicita’.
La durata della percezione consapevole del tempo e’ immodificabile. Il tempo non mi permette di modificare il passato ma mi permette di conservare i ricordi. Ogni uomo racchiude le memorie del tempo, della sua vita, cosicché’ il passato gli appartiene sotto forma di ricordo ma non di tempo, perché’ lo stesso non appartiene a nessuno, lui rappresenta l’invisibile condizione della vita. Per questo motivo quando penso al mio tempo, in questa vita, lo immagino come la cornice del quadro della mia vita.
Il maestro, compagno, consigliere o ingannatore, e’ tutto relativo a come l’uomo riesce a farne uso, e’ perciò’ importante l’utilizzo dl tempo e il traguardo probabilmente e’ rappresentato dal raggiungimento di una serenità’ pura, priva di condizionamenti. L’essere sereno spende bene il suo tempo, e probabilmente trascorrerlo con qualità’ significa averne colto l’importanza. Il problema e’ che spesso viene sottovalutato e ignorato, mal utilizzato, a causa della mancata effettiva percezione della sua presenza. E lui continua a trasformare le esperienze presenti in ricordi passati, nel nostro prossimo futuro.

Mio padre va spesso in bici e si stare chilometri sulle montagna e spesso sceglie percorsi diversi e quindi gli serve per forza un’aiuto per questo motivo utilizza un piccolo computer per biciclette, davvero comodo, ma quello che non sapevo era la quantità di tipi in commercio. In realtà infatti, i personal computer per biciclette e ciclisti possono essere classificati sia come computer multifunzione o monitor della frequenza cardiaca; molti sono stati progettati con funzioni multiple, fino a 50, mentre alcuni semplicemente monitorare la frequenza cardiaca, o hanno circa 10 funzioni chiave che aiutano a migliorare la vostra velocità o resistenza.
Storicamente parlando,m questi accessori sono stati venduti dagli anni ottanta, e hanno aiutato atleti professionisti,che li usano in gara, e anche per i ciclisti, come il mio babbo, che li utilizzano per l’uso quotidiano. Ci sono una varietà di funzioni all’interno di un ciclocomputer, ed ovviamente i prezzi variano. Molti calcolano aree comuni come velocità, distanza, frequenza cardiaca e le calorie bruciate, inoltre si adattano tramite zone cablate, e dovrebbero sempre essere venduti con le istruzioni complete per il montaggio semplice, un fai da te casalingo!
Senza fili o con i fili?
Un ciclocomputer è un monitor che viene collegato alla tua bicicletta. Quelli con i fili, potrebbero essere utili per i ciclisti regolari, non hanno bisogno di essere regolati o fissati prima usarlo, ma dovrà essere montato con cura per evitare grovigli e l’interferenza con la tua bicicletta. Al contrario il ciclocomputer wireless, senza fili, è stato progettato per essere montato come preferisci. Queste tipologie possono rivelarsi utili quando viene utilizzato come il contapassi per movimento, e sono l’ideale per i ciclisti occasionali.
I ciclocomputer senza fili possono essere rimossi, che è anche una buona caratteristica per i ciclisti urbani che possono essere a più alto rischio di furto. Il modo migliore per decidere cosa acquistare è quello di guardare a ciò che sono le vostre esigenze. Le caratteristiche più, il più costoso il dispositivo sarà. Inoltre le nuove tecnologie permettono di essere interattivi con ciclocomputer portatili, una nuova funzionalità per molti dispositivi, per cui si possono scaricare i dati sul computer per l’analisi e il monitoraggio.
Se si intende utilizzare questa funzione, allora questo potrebbe essere un grande acquisto e potrebbe contribuire a aiutare il tuo allenamento e il tuo progresso. Tieni a mente, consiglio un ciclocomputer con un montaggio incluso altrimenti dovrai acquistare un supporto che può tenerlo in modo sicuro al manubrio.

A moltissime persone non piace assolutamente stirare, È stato provato che si tratta di stirare la maggior parte delle persone vorrebbe ottenere un ottimo risultato nel miglior tempo possibile. Tuttavia con l’utilizzo dei nuovi modelli a vapore, è possibile riuscire a stirare con estrema facilità in maniera davvero molto efficace, prima cosa, perché si possono dividere i fan-e ora oggi come oggi, in commercio si ritrovano con generatore di vapore pure pressurizzati.
Le tipologie dei ferri moderni permettono di stirare con estrema facilità qualsiasi cosa ti serva, anche molto velocemente, come ho scritto sopra ci sono tipologie diverse, anche portatili. Metti caso hai una presentazione importante e porti con te il tuo abito, spesso la valigia spiegazza i vestiti, anche se impari a metterli con estrema cura, ti porti il tuo piccolo ferro da stiro e il gioco è fatto, in pochissimo tempo hai sistemato le pieghe inestetiche.
Beh, se ami stirare e hai tempo per dedicare alla stiratura puoi anche acquistare un ferro da stiro a vapore con serbatoio incorporato, e senza funzioni particolari, in modo che decidi tu la pressione del vapore e il calore del ferro, in base al tessuto, in questo caso sei un professionista e tanto di cappello, ma per me non è andata proprio così!
Il mio ferro da stiro autonomo!
Ammetto che non mi piace stirare, credo che sia anche una perdita di tempo, ma diventa indispensabile per certe circostanze e quindi devo stirare. Per semplificare il mio lavoro, ho deciso di comprare un ferro da stiro pressurizzato con caldaia separata, che in realtà funziona da solo. Dato che stiro poco, ma quando lo faccio ho il classico montone di vestiti, ho preferito acquistare una tipologia che potesse anche contenere molta acqua, in modo tale che non ho mai problemi quando devo stirare tanta roba e soprattutto non devo riempire di acqua il ferro i. Continuazione.
Inoltre, il mio ferro ha la capacità di distinguere i capi e di regolare la pressione dell’acqua e il calore in base al tessuto. Questo perché, come ti ho appena fletto no. Essendo mai sicura delle mie capacità da stiratrice, ho preferito affidar,i ad un prodotto che potesse fare quasi tutto da solo. E ovviamente dato che mio marito viaggia anche per lavoro, ho pensato di regalargli un mini ferro da stiro,che si può portate ovunque, una svolta! Ovviamente gli ho insegnato ad usarlo ed è stata una vera svolta!